Makroevolusjon: Firføttinger utviklet seg?
Oversatt via FB, herfra.
 Jeg tenkte på hvor mye hjernekraft som kreves for å programmere og lage robotene fra Boston Dynamics. I den naturlige verden, ifølge tilhengere av naturalisme, skyldtes den nødvendige koordineringen og oppfinnelsen av nye lemmer, bare tilfeldige naturlige prosesser. Det fikk meg til å se nærmere på hva vanlige vitenskapelige artikler har å si om emnet. Hvordan oppsto de første lemmene til tetrapoder? Hvilken mekanisme kreves for å vokse kroppsdeler som ben, og hvordan forklarer tilhengere av evolusjon oppkomsten av tetrapoder?
Jeg tenkte på hvor mye hjernekraft som kreves for å programmere og lage robotene fra Boston Dynamics. I den naturlige verden, ifølge tilhengere av naturalisme, skyldtes den nødvendige koordineringen og oppfinnelsen av nye lemmer, bare tilfeldige naturlige prosesser. Det fikk meg til å se nærmere på hva vanlige vitenskapelige artikler har å si om emnet. Hvordan oppsto de første lemmene til tetrapoder? Hvilken mekanisme kreves for å vokse kroppsdeler som ben, og hvordan forklarer tilhengere av evolusjon oppkomsten av tetrapoder?
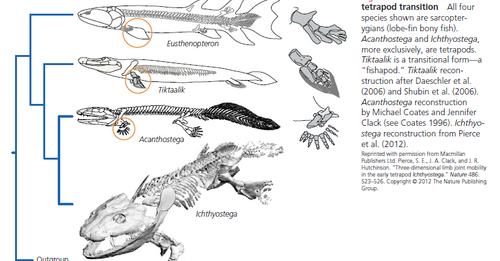
Ifølge tilhengere av evolusjon oppsto tetrapoder fra en avstamning av fisk. Denne typen dramatisk endring over tid kalles makroevolusjon.
Overgangen fra liv i vann til liv på land ville ha nødvendiggjort dramatiske strukturelle endringer av hele kroppen for å motstå de økte effektene av tyngdekraften, blant andre nye krav.
Mange aspekter av tetrapod-opprinnelse forblir unnvikende. Dens antatte utvikling har skapt stor interesse, men de tidligste fasene av historien deres er dårlig forstått. Nyere studier har stilt spørsmål ved lenge aksepterte hypoteser om opprinnelsen til pentadactyl-lemmet, fylogenien til tetrapoder og miljøet der de første tetrapodene levde.
De ‘tidligste’ kjente tetrapodene med føtter og ben antas nå å ha vært vannlevende dyr; tilhengere av evolusjon hevder derfor at føtter og ben utviklet seg i et grunt vannmiljø og først senere ble valgt for bruk på land.
Bilde 1. Forekomster av firføtttinger (tetrapoder)
De fleste diskusjoner om emnet konsentrerer seg om å belyse om fossilregistreringen tillater å finne overgangsformer som tillater å antyde en overgang fra vann til land. Ikke bare er det store hull, men ideen bærer store problemer konseptuelt, og som helhet.
 Dessuten, som Behe fint forklarte: For å si at en eller annen funksjon er forstått, må alle relevante trinn i prosessen belyses. De relevante trinnene i biologiske prosesser skjer til syvende og sist på molekylært nivå, så en tilfredsstillende forklaring på et biologisk fenomen som oppkomsten av tetrapoder må inkludere hvordan overgangen skjedde på en molekylær forklaring. Det er ikke lenger tilstrekkelig for en "evolusjonær forklaring" av denne kraften å påberope seg de anatomiske strukturene til hele øyne, slik Darwin gjorde på 1800-tallet og som de fleste popularisatorer av evolusjon fortsetter å gjøre i dag. Anatomi er rett og slett irrelevant. Det samme er fossilregisteret. Det spiller ingen rolle om fossilene stemmer overens med evolusjonsteorien eller ikke, like mye som det betydde noe i fysikk at Newtons teori var i samsvar med hverdagserfaring. Fossilregisteret har ingenting å fortelle oss om for eksempel om eller hvordan interaksjonene mellom 11-cis-retinal og rhodopsin, transducin og fosfodiesterase kunne ha utviklet seg trinnvis. Heller ikke mønstrene for biogeografi spiller noen rolle, eller populasjonsgenetikk, eller forklaringene som evolusjonsteorien har gitt for rudimentære organer eller artsoverflod.
Dessuten, som Behe fint forklarte: For å si at en eller annen funksjon er forstått, må alle relevante trinn i prosessen belyses. De relevante trinnene i biologiske prosesser skjer til syvende og sist på molekylært nivå, så en tilfredsstillende forklaring på et biologisk fenomen som oppkomsten av tetrapoder må inkludere hvordan overgangen skjedde på en molekylær forklaring. Det er ikke lenger tilstrekkelig for en "evolusjonær forklaring" av denne kraften å påberope seg de anatomiske strukturene til hele øyne, slik Darwin gjorde på 1800-tallet og som de fleste popularisatorer av evolusjon fortsetter å gjøre i dag. Anatomi er rett og slett irrelevant. Det samme er fossilregisteret. Det spiller ingen rolle om fossilene stemmer overens med evolusjonsteorien eller ikke, like mye som det betydde noe i fysikk at Newtons teori var i samsvar med hverdagserfaring. Fossilregisteret har ingenting å fortelle oss om for eksempel om eller hvordan interaksjonene mellom 11-cis-retinal og rhodopsin, transducin og fosfodiesterase kunne ha utviklet seg trinnvis. Heller ikke mønstrene for biogeografi spiller noen rolle, eller populasjonsgenetikk, eller forklaringene som evolusjonsteorien har gitt for rudimentære organer eller artsoverflod.
Bilde 2. Forsøk på oppsett anatomisk utvikling
Så i stedet for å holde oss til anatomiske sammenligninger av fossiler som kan ha en viss likhet som kan tolkes som mellomprodukter og utvikling av tetrapodlemmer fra fiskefinner, la oss prøve å belyse hvor betydelig det funksjonelle og morfologiske skiftet var med tanke på de underliggende genetiske mekanismene . Fossilregisteret gir innsikt i antatte morfologiske endringer. For å forstå de underliggende mekanismene, må vi imidlertid kikke inn i de genregulerende nettverkene til levende virveldyr.
Oppstår nye anatomiske strukturer de novo, eller utvikler de seg fra eksisterende strukturer? Fremskritt innen utviklingsgenetikk, paleontologi og evolusjonær utviklingsbiologi har nylig visstnok kastet lys over opprinnelsen til strukturene som fascinerte Charles Darwin mest, inkludert tetrapod-lemmer. I følge tilhengere av evolusjon oppsto strukturer ved modifisering av eksisterende genetiske reguleringskretser.
Grassos kommentar: Det genetiske programmet instruerer hvordan man lager nye strukturer, men det programmet må være nøyaktig programmert, og de genetiske reguleringskretsene må også programmeres. Det vil si at to separate programmer må dukke opp, det vil si 1. programmet som definerer den fysiske formen og strukturen, og 2. programmet som instruerer hvor man finner den genetiske informasjonen i genomet, og når den skal uttrykkes under utviklingen, at det er i riktig rekkefølge. Det er forskjellige lag med informasjon, som må eksistere fullt utviklet for å lage de nye anatomiske delene det gjelder.
Instruksjonene som kontrollerer når og hvor et gen uttrykkes er skrevet i sekvensen av DNA-baser som ligger i den regulatoriske regionen av genet. Disse instruksjonene er skrevet på et språk som ofte kalles "genreguleringskoden". Denne koden leses og tolkes av proteiner kalt transkripsjonsfaktorer som binder seg til spesifikke sekvenser av DNA (eller 'DNA-ord') og øker eller reduserer genuttrykk. Endringer i genuttrykk mellom arter kan derfor skyldes endringer i transkripsjonsfaktorene og/eller endringer i instruksjonene innenfor de regulatoriske områdene til spesifikke gener.
 For at kommunikasjon skal skje, kreves 1. Sekvensen av DNA-baser lokalisert i den regulatoriske regionen av genet, og 2. transkripsjonsfaktorer som leser koden. Hvis en av begge mangler, svikter kommunikasjonen, genet som skal uttrykkes, kan ikke påtreffes, og hele prosedyren for genuttrykk mislykkes. Dette er et irreduserbart komplekst system. Den genregulerende koden kunne heller ikke oppstå på en trinnvis måte, siden hvis det var tilfelle, har koden bare den rette betydningen hvis den er ferdig utviklet. Det er et par excellence eksempel på intelligent design.
For at kommunikasjon skal skje, kreves 1. Sekvensen av DNA-baser lokalisert i den regulatoriske regionen av genet, og 2. transkripsjonsfaktorer som leser koden. Hvis en av begge mangler, svikter kommunikasjonen, genet som skal uttrykkes, kan ikke påtreffes, og hele prosedyren for genuttrykk mislykkes. Dette er et irreduserbart komplekst system. Den genregulerende koden kunne heller ikke oppstå på en trinnvis måte, siden hvis det var tilfelle, har koden bare den rette betydningen hvis den er ferdig utviklet. Det er et par excellence eksempel på intelligent design.
https://reasonandscience.catsboard.com/t2220-shannons-theory-of-information?fbclid=IwAR1ds7w_9jrTVrt1rpEjETl1uxvSsSvzIOFtTzdFc_9CtfIoVK9rNDkN9v8
Under utviklingen av vertebratens lemmer transkriberes Hoxd-gener i to temporale faser; en tidlig bølge kontrollerer vekst og polaritet opp til underarmen og en sen bølge mønstrer sifrene. I denne utgaven av Developmental Cell rapporterer Tarchini og Duboule (2006) at to motsatte regulatoriske moduler styrer tidlig kolinær ekspresjon av Hoxd-gener.
Spørsmål: hvordan kunne naturlige mekanismer ha programmert og styrt de riktige tidsfasene av gentranskripsjon av de riktige genene, og tidlig bølgekontroll? Videre utvikler lemmene seg på rett sted, det kreves riktige koordinater og posisjonsinformasjon, og de kan utvikle seg hvor som helst på kroppen. Fant naturlige mekanismer ut om det rette stedet ved prøving og feiling? Det var myriader av posisjoner mulig å legge til lem. Hvordan kunne riktig og presis koordinering av aksial posisjon oppnås ved mutasjoner?
Bilde 3. Regulering krever programmering (Grasso)
Problemet er at naturen har for mange alternativer og kunne ikke sortere dem alle, uten design. Naturlige mekanismer er for uspesifikke til å bestemme noe bestemt utfall. Mutasjon og naturlig seleksjon kan teoretisk sett danne et nytt komplekst morfologisk trekk som et ben eller et lem med riktig størrelse og form, og sørge for å finne ut det rette kroppsstedet for å vokse dem, men det kan også produsere alle slags andre nye kroppsformer , og vokse og feste dem hvor som helst på kroppen, hvorav de fleste ikke har noen biologiske fordeler eller mest sannsynlig er skadelige for organismen. Naturlige mekanismer har ingen begrensninger, de kan produsere enhver form for nyhet. Det er imidlertid den typen frihet som gjør det ekstremt usannsynlig at bare naturlige utviklinger gir nye spesifikke evolusjonære arrangementer som er fordelaktige for organismen. Naturen ville måtte arrangere nesten et uendelig antall forsøk og feil før man fikk en ny positiv ordning. Siden det ville blitt en svært usannsynlig hendelse, er design en bedre forklaring.
Når jeg gikk gjennom flere vitenskapelige artikler, har jeg ikke kommet over én av dem, som var i stand til å gi en detaljert beskrivelse av nøyaktig hvordan den morfologiske overgangen kunne ha skjedd gjennom evolusjon.
Noen biologer har også sett for seg spesielle mutasjoner i regulatoriske homeobox- eller "Hox"-gener, der enkle mutasjoner kan være i stand til å gjøre store utviklingsendringer i en organisme som kan forårsake en radikalt annen fenotype. Imidlertid gjør manipulering av "Hox"-gener lite for å løse problemet med å generere nye funksjonelle biostrukturer, for å gjøre store endringer i fenotype er sjelden fordelaktig. Hox-genmutasjoner kan være en enklere mekanisme for å generere store endringer, men de slipper heller ikke unna problemet med det "håpefulle monsteret":" Ulempen for forskere er at  naturens skarpsindige økonomi skjuler enorm kompleksitet. Forskere finner bevis på at Hox gener og ikke-Hox homeobox-genene er ikke uavhengige agenter, men medlemmer av enorme genetiske nettverk som forbinder hundrevis, kanskje tusenvis, av andre gener. Endre én komponent, og utallige andre vil endre seg også - og ikke nødvendigvis til det bedre. Dermed vil drømmer om å fikle med naturens verktøykasse for å bringe til live det forskerne kaller et "håpefullt monster" - for eksempel en fisk med føtter - sannsynligvis forbli unnvikende." Videre glemmer mange biologer når de påkaller Hox-genmutasjoner at Hox-gener bare kan gjenopprette -arranger deler som allerede er der - de kan ikke lage virkelig nye strukturer.
naturens skarpsindige økonomi skjuler enorm kompleksitet. Forskere finner bevis på at Hox gener og ikke-Hox homeobox-genene er ikke uavhengige agenter, men medlemmer av enorme genetiske nettverk som forbinder hundrevis, kanskje tusenvis, av andre gener. Endre én komponent, og utallige andre vil endre seg også - og ikke nødvendigvis til det bedre. Dermed vil drømmer om å fikle med naturens verktøykasse for å bringe til live det forskerne kaller et "håpefullt monster" - for eksempel en fisk med føtter - sannsynligvis forbli unnvikende." Videre glemmer mange biologer når de påkaller Hox-genmutasjoner at Hox-gener bare kan gjenopprette -arranger deler som allerede er der - de kan ikke lage virkelig nye strukturer.
Bilde 4. dGRN bananflue
Casey Luskin: Hox-mutasjoner vil aldri skape nye "kroppsdelgener", og kan dermed ikke legge til virkelig nye fenotypiske funksjoner i genomet, og i beste fall sitter vi igjen med dilemmaene knyttet til "pre-adaptation". Størstedelen av evolusjonær endring må skje gjennom utvikling av nye "kroppsdelgener", noe Hox-mutasjoner ikke kan gjøre. En anmelder i Nature gjenkjenner dette faktum: "Schwartz ignorerer det faktum at homeobox-gener er selektorgener. De kan ikke gjøre noe hvis genene som reguleres av dem ikke er der. Det er disse genene som i detalj spesifiserer den adaptive strukturen til organene. Vær sikker på, at å slå på et homeobox-gen på feil sted kan føre til utseendet til et ektopisk organ, men bare hvis genene for det organet er tilstede i samme individ. Det er helt feil å antyde at et øye kan være produsert av en makromutasjon når det aldri var noe øye tilstede i avstamningen før.
Darwins tvil, s.239
HVA MED HOX-GENER? Hox (eller homeotiske) gener regulerer uttrykket av andre proteinkodende gener under prosessen med dyreutvikling. Noen biologer har sammenlignet dem med dirigenten for et orkester som spiller rollen som å koordinere bidragene til spillerne. Og fordi Hox-gener påvirker så mange andre gener, tror mange evo-devo-forkjempere at mutasjoner i disse genene kan generere store endringer i form.
Bilde 5. Mutert hox-gen ('antenne')
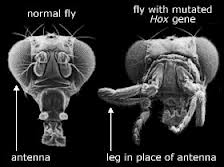 Men kan mutasjoner i Hox-gener transformere én form for dyreliv - én kroppsplan - til en annen? Det er flere grunner til å tvile på at de kan.
Men kan mutasjoner i Hox-gener transformere én form for dyreliv - én kroppsplan - til en annen? Det er flere grunner til å tvile på at de kan.
For det første, nettopp fordi Hox-gener koordinerer uttrykket av så mange andre forskjellige gener, har eksperimentelt genererte mutasjoner i Hox-gener vist seg å være skadelige. i bananfluer "forårsaker de fleste mutasjoner i homeotiske [Hox] gener dødelige fødselsskader." I andre tilfeller er den resulterende Hox-mutante fenotypen, selv om den er levedyktig på kort sikt, ikke desto mindre markant mindre egnet enn villtypen. For eksempel, ved å mutere et Hox-gen i en fruktflue, har biologer produsert den dramatiske Antennapedia-mutanten, en ulykkelig flue med ben som vokser ut av hodet der antennene skal være.
https://reasonandscience.catsboard.com/t2077-hox-genes?fbclid=IwAR2NrvCo3MtY4nDTZZVobKbdjHSBRoW7rUfr3AhcJph9SToRvXQPYH8DT98
Oversettelse og bilder ved Asbjørn E. Lund